Zebrafish Evolution

The zebrafish, Danio rerio, is a shimmering, striped creature that has become one of the world's most important biomedical models, primarily thanks to the pioneering work of George Streisinger and his colleagues in the 1970s and 1980s. Its rapid development, external embryos, and genetic tractability propelled it to the top tier of NIH-funded research, providing a sequenced genome and tools for genetic manipulation. Yet, for all the knowledge gained from this 'system,' the story of the organism itself—its ecological background and evolutionary history—is equally compelling and increasingly vital for grounding our scientific inferences. Understanding where the zebrafish came from helps us understand what it means when we apply our findings to other vertebrates, including humans.
# Native Environment
The journey of Danio rerio begins in the freshwater habitats of South Asia, where it was first formally described by the Scottish physician Francis Hamilton in the state of Bihar, northeastern India. Its current confirmed range stretches north into Pakistan and Nepal, and east into Bangladesh and possibly Myanmar. These fish are not found in the main channels of massive rivers like the Ganges, but rather thrive in specific, less extreme environments: shallow, slow-moving streams, and particularly the still pools that form near stream margins during the monsoon season.
Wild zebrafish are highly adaptable generalists when it comes to abiotic factors. They inhabit areas ranging from elevations of about 8 meters up to nearly 1,600 meters above sea level, tolerating water temperatures that swing widely, sometimes between and , and experiencing pH levels from nearly neutral to somewhat basic (5.9 to 9.8). This broad tolerance across varying physical conditions suggests that the high genetic diversity observed in wild populations is not merely historical noise; rather, it represents the raw, adaptive material that allows local populations to maintain fitness across different hydrological basins and seasonal shifts, an essential context for studying local adaptation. Their habitat is often shallow, with sandy or muddy bottoms, featuring submerged or overhanging vegetation that provides cover. Because they have shared space with humans for millennia, they have also successfully colonized man-made environments like rice paddies and drainage ditches, though they consequently suffer from pollution and habitat loss.
As omnivores, their natural diet is varied, consisting of small crustaceans, insects, zooplankton, algae, and assorted detritus, a pattern of eating that differs significantly from the consistent, protein-rich laboratory diet fed to research specimens. This ecological niche places them in constant interaction with a host of other organisms. They compete for resources with other small minnows like Esomus and Puntius, and they are prey for larger fish, birds, and even dragonfly larvae, especially during their early, highly vulnerable egg and larval stages.
# A Fish Family

To place the zebrafish in evolutionary time, we must look at its taxonomic home. Danio rerio belongs to the family Danionidae within the order Cypriniformes, a lineage that represents the most species-rich group of all vertebrates, the teleosts (bony fishes). The subfamily Danioninae to which it belongs shows fascinating morphological extremes. While the zebrafish itself grows to a modest 4–5 cm, its relatives include species like Devario aequipinnatus (the giant danio), reaching about 13 cm, and others that represent some of the smallest vertebrates known, such as Danionella and Paedocypris, which mature at only 1 to 1.5 cm in a larval-like form. This extreme miniaturization appears to have evolved independently in at least two different lineages within the subfamily.
Phylogenetic studies aiming to precisely map the relationships within the Danio genus are complex, partly because gene flow and hybridization have occurred between lineages, complicating the clear demarcation of species boundaries based on simple genetic trees. For instance, resolving the closest relatives to D. rerio has been difficult, though Danio aesculapii has emerged as a candidate sister species based on extensive genomic sampling. The study of these relatives provides a crucial comparative backdrop; for example, if a trait like spinal development is conserved across the Danio genus, its mechanism is likely evolutionarily deep-seated, whereas a trait that differs wildly, like adult color pattern, may show a more recent divergence in underlying mechanisms.

# Pigment Pattern Evolution
Perhaps the most obvious evolutionary feature of D. rerio is its namesake pattern: five uniform, pigmented, horizontal blue stripes on a laterally compressed body. This distinctive adult striping contrasts sharply with the diversity found across its relatives in Danio, where patterns include vertical bars, spots, or reduced stripes. When zebrafish are crossed with other Danio species, the striped pattern of the zebrafish often dominates in the hybrids, suggesting that the alleles responsible for this look are frequently dominant over those in related species.
The development of these stripes is a complex cellular process, involving interactions between pigment cell lineages—specifically melanophores (black), iridophores (reflective), and xanthophores (yellow)—and is known to depend on positional cues and thyroid hormone signaling. Interestingly, the larval pigment patterns across different Danio species are nearly identical. One compelling hypothesis is that this conserved pattern evolved early in the lineage's history to serve as a protective UV shield for developing vital structures, like the central nervous system and gonads, as they developed in shallow, sunny waters. It is remarkable that the mechanisms to produce this ancestral pattern remain accessible, while the adult pattern is the result of a more recent, hormone-regulated refinement involving differential recruitment of pigment stem cell lineages. This points to the evolution of regulatory elements—the genetic switches that control when and where genes are expressed—as key drivers of species differentiation. For example, the uniform pattern seen in D. albolineatus is linked to cis regulatory changes that alter the timing of pigment cell differentiation.
# Lab Adaptation Versus Wild Diversity
The genetic comparison between laboratory-maintained strains and their wild progenitors offers one of the most insightful evolutionary case studies available for any model organism. The founding stocks of lab zebrafish were derived from a limited number of wild fish, often acquired via the pet trade, meaning they experienced intense founder effects and subsequent selective pressures unique to laboratory life—sometimes intentional selection for specific traits, and other times relaxed selection on traits important only in the wild, such as predator avoidance.
The resulting genetic chasm is vast. While the sequencing of the reference genome revealed about 7 million Single Nucleotide Polymorphisms (SNPs) between two lab strains, a single wild zebrafish can harbor over 5 million SNPs within its own genome. Comparing this to humans, where thousands of sequenced genomes yield a total of only 38 million SNPs, suggests that SNPs are approximately four times more frequent in zebrafish than in humans, relative to genome size. Furthermore, Copy Number Variants (CNVs) are about 1.5 times more prevalent in zebrafish. This incredible genetic diversity in the wild underpins their ecological adaptability, but it presents a significant challenge in the lab: genetic background effects can confound results unless rigorously controlled.
A particularly striking example of domestication-driven evolution involves sex determination. Many fish species use clear XY or WZ systems. For years, researchers struggled to find a consistent sex-determining system in lab zebrafish, as regions associated with sex differed between studies. The analysis of wild populations revealed the reason: wild D. rerio possess a major sex determinant () on Chromosome 4, a system similar to that in birds, but this determinant has been lost in lab strains. The current, often complex, sex determination in lab strains is likely the result of genetic drift and selection acting on multiple, smaller effect loci after the primary determinant was lost—a classic evolutionary story where human intervention inadvertently created a new, functionally distinct population.

# Spine Development and Deep Time
The evolutionary relevance of zebrafish extends far beyond the differences between their lab and wild cousins; their developmental toolkit holds clues to vertebrate history. The way bony fish, or teleosts, construct their spines differs fundamentally from land animals like mammals. In teleosts, the central structure is the notochord, a tube that patterns the surrounding cells to form articulated vertebrae. In contrast, the notochord in human embryos gives rise to the intervertebral discs found between our vertebrae.
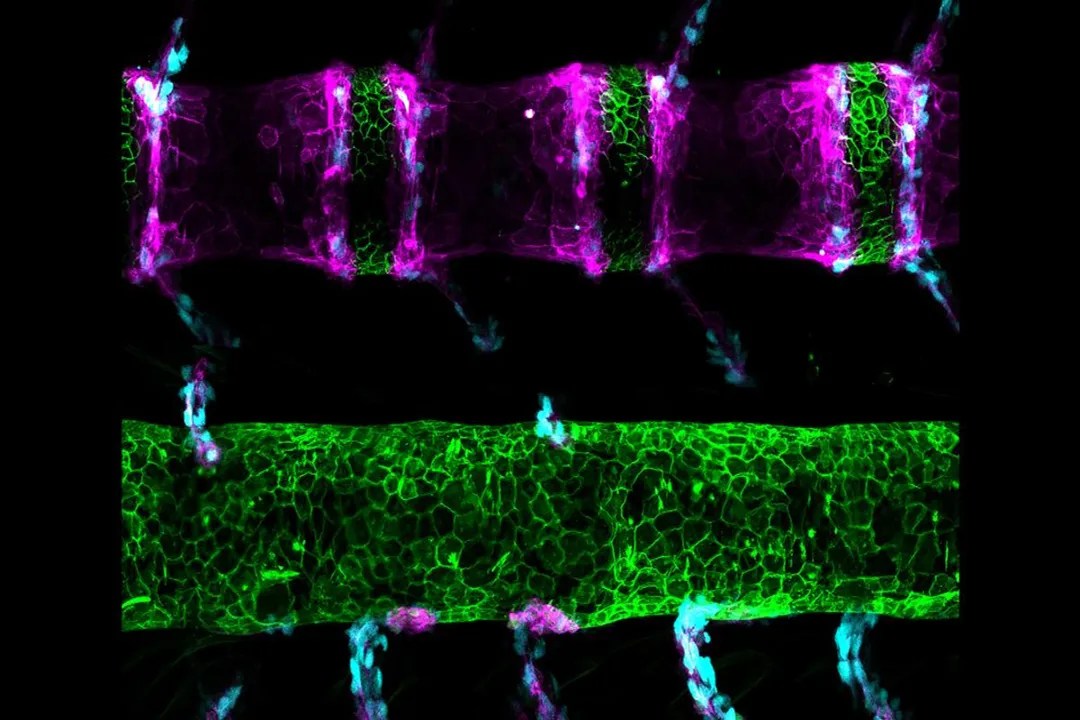
This evolutionary split was illuminated by a chance finding in a mutant line named spondo (Greek for spine), caused by a single-letter change in a teleost-specific gene that regulates notochord patterning. The spondo mutant fish developed a "tortured looking spine" with vertebrae cleaved in half, closely resembling fossil specimens of ancestral fish. This is a profound example of evolutionary developmental biology (EvoDevo) in action. The research indicated that the genetic recipe for the ancestral spine is still present in the zebrafish genome, simply hidden by a newer regulatory system. The single DNA change in spondo effectively reverted the developmental program to a more ancient state. This tiny genetic difference may mark the point where the developmental pathways leading to our mammalian spines parted ways with those of our teleost ancestors over vast evolutionary time. The observation shows that developmental pathways are not erased but rather layered, and that a small genetic alteration can reveal previously hidden evolutionary potential—a testament to the cryptic depth of the vertebrate genome.
# Bridging Homology and Analogy
As zebrafish research increasingly intersects with human medicine, the question of evolutionary homology becomes central to justifying the model’s use. Similarity between species can arise from shared ancestry (homology) or convergent evolution due to similar selective pressures (analogy). For discoveries to be translationally relevant, we require similarity in the underlying mechanisms—that is, homology.
At the genetic level, homology is relatively straightforward to establish; zebrafish share about 70% of their genes with humans, and synteny (the order of genes on chromosomes) shows conserved blocks. However, as the level of analysis moves up to function and behavior, things become murkier. While teleosts and mammals share homologous brain structures—for instance, the zebrafish lateral pallium is homologous to the mammalian hippocampus, both implicated in relational learning—their physical placement differs due to distinct brain development processes (eversion in fish versus evagination in mammals).
Behavioral similarity, such as the high sociality displayed by both zebrafish (shoaling) and humans, is even harder to interpret evolutionarily. While zebrafish exhibit behaviors like shoaling, which likely evolved for predator protection or foraging, researchers must be mindful of ethological relevance—ensuring lab observations reflect natural behavior. The fact that zebrafish can exhibit strong antipredator responses simply from being handled by a researcher, or that their natural foraging habits (small, sporadic meals) make classic food-reward learning tasks difficult, shows how the artificial lab context can distort the output of the brain. To avoid studying analogy when we need homology, one must deeply characterize the natural stimuli and motivations that shape behavior in the wild.
By studying the ecology, population genetics, and phylogenetic relationships of the zebrafish—from the high genetic diversity in the Ganges-Brahmaputra basin to the specialized mechanisms that determine spine structure—we gain vital context. This understanding allows us to better interpret the conserved genetic pathways that drive development and disease, while also refining experimental design to ensure that the marvelous 'system' accurately reflects the evolutionary history of the 'organism'. The zebrafish, a creature adapted to monsoon pools, continues to offer windows not only into vertebrate development but into the very deep evolutionary architecture that shapes life today.
Related Questions
#Citations
The Natural History of Model Organisms: Advancing biology through ...
Advancing biology through a deeper understanding of zebrafish ...
Zebrafish - Wikipedia
EvoDevo in Zebrafish: Bridging Evolution, Development, and Drug ...
Graduate Student's Mutant Zebrafish Reveals Evolutionary History